
Der Insulinrezeptor sieht keinen Zucker: Wie Zellen ein Hormonsignal in Transportarbeit übersetzen
- Benjamin Metzig
- 7. Juni
- 5 Min. Lesezeit

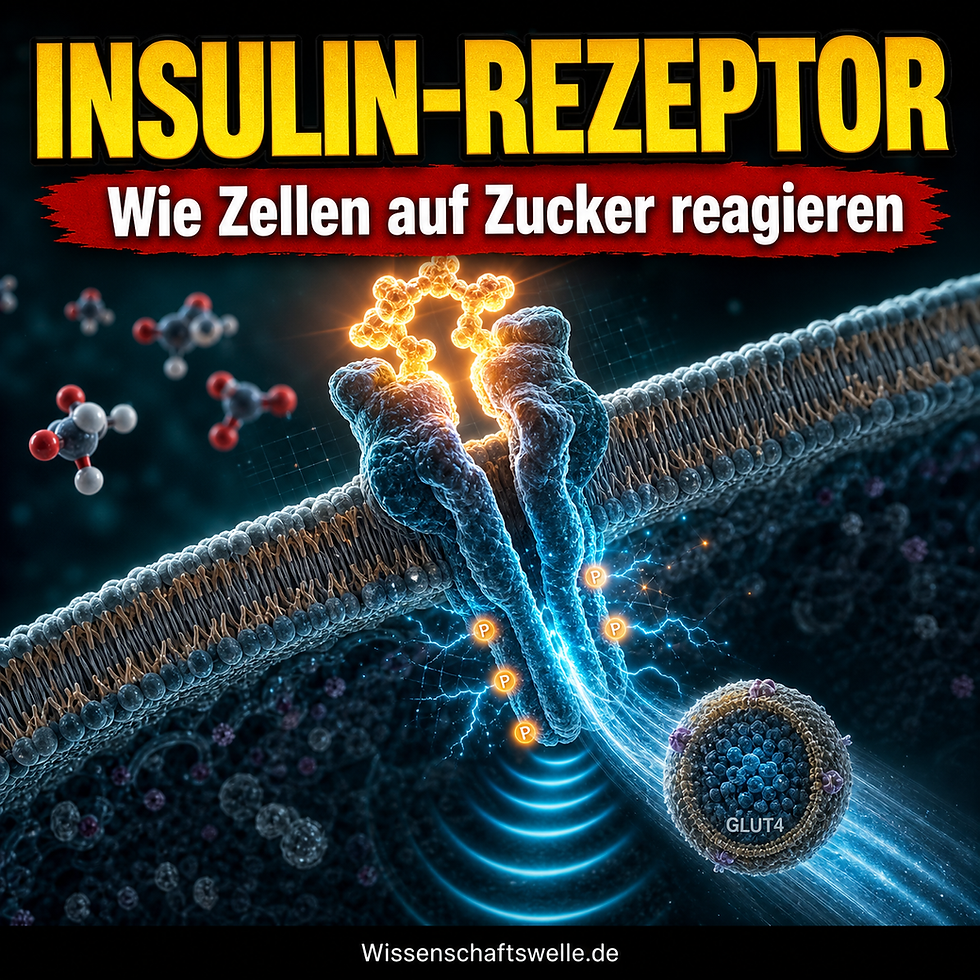
Wenn nach einer Mahlzeit der Blutzucker steigt, klingt es fast selbstverständlich zu sagen: Jetzt müssen die Zellen den Zucker erkennen und aufnehmen. Biochemisch passiert aber etwas anderes. Die meisten Zellen reagieren nicht direkt auf Glukose, sondern auf ein Hormon, das deren Ankunft meldet. Der entscheidende Übersetzer heißt Insulinrezeptor. Er sitzt bereits als vorgeformter Signalapparat in der Zellmembran und macht aus einer äußeren Bindung eine ganze Kette innerer Entscheidungen.
Kernaussagen
Der Insulinrezeptor ist kein Zuckersensor, sondern ein Rezeptor für das Hormon Insulin.
Seine eigentliche Schaltarbeit beginnt mit Tyrosin-Phosphorylierungen und Adapterproteinen wie IRS, nicht mit Glukose selbst.
In Muskel- und Fettzellen führt das Signal vor allem dazu, dass GLUT4-Transporter an die Zelloberfläche gebracht werden.
In der Leber geht es weniger um GLUT4 als darum, Glukoseproduktion, Glykogenspeicherung und Stoffwechselprogramme neu zu ordnen.
Insulinresistenz bedeutet meist keine einzelne kaputte Stelle, sondern eine gestörte Übersetzung an mehreren Schritten derselben Signalkette.
Der Rezeptor wartet schon an der Membran
Der Insulinrezeptor gehört zur Familie der Rezeptor-Tyrosinkinasen, ist aber innerhalb dieser Familie ein Sonderfall: Er muss nicht erst durch Ligandenbindung zu einem Dimer zusammenkommen, sondern liegt bereits als verknüpfter α2β2-Komplex vor. Das beschreibt das Endotext-Kapitel zum Insulinrezeptor und seinem Signalnetzwerk sehr präzise. Zwei äußere α-Untereinheiten binden Insulin, zwei β-Untereinheiten durchspannen die Membran und tragen innen die Kinasedomänen.
Gerade diese Vororganisation ist wichtig. Der Rezeptor muss nicht erst gefunden und zusammengesetzt werden, sondern steht gewissermaßen scharf an der Oberfläche. Neuere Strukturarbeiten zeigen zudem, dass Aktivierung kein simples Ein-Aus-Prinzip ist. Die cryo-EM-Studie von Uchikawa und Kolleginnen und die spätere Arbeit zur synergistischen Aktivierung über zwei Bindungsstellen legen nahe, dass partielle und volle Aktivierung strukturell unterscheidbar sind. Mehrere Insulinmoleküle können an verschiedene Stellen des Rezeptors koppeln und so unterschiedlich stabile Aktivierungszustände erzeugen.
Das ist mehr als ein molekulares Detail. Es erklärt, warum der Insulinrezeptor nicht wie ein mechanischer Schalter funktioniert, sondern wie ein fein abgestimmtes Empfangsgerät. Schon auf der Ebene der Bindung steckt also Abstufung.
Aus Bindung wird eine Befehlskette
Sobald Insulin bindet, phosphoryliert der Rezeptor bestimmte Tyrosinreste an sich selbst. Diese Autophosphorylierung verändert nicht nur seine Aktivität, sondern schafft auch Dockingstellen für Adapterproteine, vor allem für IRS-Proteine. Von dort verzweigt sich das Signal in mehrere Richtungen. Die zentrale metabolische Route läuft über PI3K, PIP3 und Akt; parallel existieren weitere Äste, etwa über MAP-Kinasen, die stärker mit Wachstum und Genregulation zu tun haben. Einen guten Überblick über diese Verzweigungen liefert die Übersichtsarbeit Biochemical and cellular properties of insulin receptor signalling.
Für die Stoffwechselwirkung ist entscheidend, dass der Rezeptor nicht einfach "mehr Zucker rein" befiehlt. Er ändert, welche Proteine wo aktiv sind, welche Transportvesikel mobilisiert werden und welche Gene eher hoch- oder herunterreguliert werden. Insulin ist deshalb kein einzelner Befehl, sondern ein Zustandswechsel.
Dass solche Signalwege nur funktionieren, wenn auf Phosphorylierung auch präzise Gegenregulation folgt, zeigt auf einer allgemeineren Ebene auch der Wissenschaftswelle-Text über Phosphatasen als Gegenspieler der Kinasen. Ein Signalweg ist nur dann steuerbar, wenn er auch wieder sauber zurückgenommen werden kann.
GLUT4 ist der spektakuläre, aber nicht einzige Effekt
Das bekannteste Bild der Insulinwirkung stammt aus Muskel- und Fettzellen: Intrazelluläre Vesikel mit dem Glukosetransporter GLUT4 werden an die Zelloberfläche gebracht. Genau dadurch steigt die Glukoseaufnahme nach einer Mahlzeit stark an. Die Fachübersicht GLUT4 On the move zeigt, dass das kein einmaliger Transportakt ist, sondern eine zeitlich gestufte Choreografie. Zuerst kommt ein schneller Schub aus einem reservierten Vesikelpool, danach ein anhaltendes Recycling über größere zelluläre Umlaufbahnen.
Das ist biochemisch elegant, weil die Zelle ihre Oberfläche nicht dauerhaft mit Transportern vollstellt. Sie hält Kapazität in Bereitschaft und setzt sie bei Bedarf frei. So wird aus einem Hormonsignal in Sekunden eine veränderte Membran.
Gerade hier wird verständlich, warum Blutzuckerregulation nicht bloß eine Sache der Bauchspeicheldrüse ist. Das Hormon mag aus dem Pankreas kommen, die eigentliche Senkung des Blutglukosespiegels hängt aber davon ab, ob Millionen Zellen ihre Transportlogistik tatsächlich umstellen.
Die Leber öffnet keine GLUT4-Schleusen
Wer Insulin nur über GLUT4 erklärt, verpasst die Leber. Sie reagiert auf Insulin anders als Muskel und Fett. Hepatozyten nutzen vor allem GLUT2, also keinen insulinabhängigen GLUT4-Schalter. Die Insulinwirkung besteht hier stärker darin, die Glukosefreisetzung zu drosseln, Glykogensynthese zu fördern und die Expression gluconeogener Programme zu bremsen. Genau diese gewebespezifische Perspektive arbeitet die große Übersicht Mechanisms of Insulin Action and Insulin Resistance heraus.
Das heißt: In Muskelzellen organisiert Insulin vor allem Import. In der Leber organisiert es vor allem Zurückhaltung. Der gleiche Rezeptor startet also nicht überall dieselbe sichtbare Antwort, sondern setzt ein gemeinsames Signal in gewebetypische Arbeit um.
Diese Unterscheidung ist wichtig, weil sie viele Missverständnisse auflöst. Wenn vom "Zucker aus dem Blut holen" die Rede ist, stimmt das für Muskel und Fett weit eher als für die Leber. Dort geht es stärker darum, weniger neuen Zucker abzugeben. Stoffwechsel ist keine Standbildlogik. Wer das vertiefen will, findet im Wissenschaftswelle-Text Fluxomics: Warum der Stoffwechsel kein Foto ist eine gute Anschlussstelle.
Dieselbe Nachricht, verschiedene Gewebe
Am Ende ist der Insulinrezeptor nur dann verständlich, wenn man ihn nicht isoliert betrachtet. Er sitzt in einem System, in dem Gewebe verschiedene Pflichten übernehmen. Muskel ist der große Verbraucher, Fettgewebe puffert und speichert, die Leber stabilisiert das Angebot zwischen Nahrungszustand und Fasten. Dass der Körper je nach Schlaf, Bewegung, Mahlzeitenrhythmus und Energieverfügbarkeit unterschiedlich auf diese Signale antwortet, ist eine breitere Frage der metabolischen Flexibilität.
Insulin ist darin keine moralische Figur, weder Held noch Feind. Es ist ein Koordinationssignal. Gerade deshalb ist die verbreitete Alltagserzählung vom "bösen Insulin" biochemisch zu grob. Ohne dieses Signal ließe sich die Arbeitsteilung zwischen Geweben gar nicht sauber organisieren.
Wenn die Übersetzung stockt
Insulinresistenz bedeutet deshalb auch nicht automatisch, dass Zellen Insulin gar nicht mehr wahrnehmen. Häufiger ist die Antwort abgeschwächt, verschoben oder nur in bestimmten Geweben und Signalästen gestört. Bereits das NIDDK erklärt in seiner Übersicht zu Insulinresistenz und Prädiabetes, dass der Körper dann mehr Insulin benötigt, um dieselbe Wirkung zu erzielen.
Auf molekularer Ebene können solche Störungen an verschiedenen Stellen ansetzen: an der Rezeptoraktivierung selbst, bei IRS-Proteinen, in nachgeschalteten Lipid- und Kinasesignalen oder in einer Umgebung aus Überernährung, Entzündung und veränderter Fettspeicherung, die die Übersetzung des Signals unpräzise macht. Genau deshalb wirkt Insulinresistenz in der Praxis so widersprüchlich. Manche Prozesse reagieren noch, andere schon deutlich weniger.
Auch hier hilft eine systemische Perspektive mehr als eine eindimensionale. Der ältere Wissenschaftswelle-Beitrag Hungerhormone verstehen zeigt das auf der Ebene von Appetit und Sättigung: Insulin ist nie nur ein Blutzuckerwerkzeug, sondern immer Teil eines größeren hormonellen Gesprächs.
Was der Insulinrezeptor eigentlich leistet
Der eigentliche Clou des Insulinrezeptors liegt nicht darin, Zucker zu erkennen. Er leistet etwas Raffinierteres: Er übersetzt die Information "Nährstoffe sind angekommen" in geordnetes Zellverhalten. Mal heißt das, Transporter an die Oberfläche zu schicken. Mal heißt es, Glukoseproduktion zu drosseln. Mal heißt es, Speicherprogramme zu aktivieren und konkurrierende Prozesse herunterzufahren.
Blutzuckerregulation ist deshalb keine direkte Begegnung von Zelle und Zucker, sondern das Ergebnis einer molekularen Verwaltungskette. Der Insulinrezeptor steht am Anfang dieser Kette. Erst wenn man ihn so betrachtet, wird verständlich, warum ein einziges Hormon gleichzeitig schnell, fein abgestuft und gewebespezifisch wirken kann.
Autorenprofil
Benjamin Metzig ist Gründer, Autor und redaktionell Verantwortlicher von Wissenschaftswelle.de. Wissenschaftswelle ist ein persönlich geführtes redaktionelles Wissensprojekt, das komplexe Themen aus unterschiedlichen Fachbereichen sorgfältig recherchiert, strukturiert und verständlich aufbereitet. Moderne Recherche-, Analyse- und KI-Werkzeuge dienen dabei als Unterstützung, während Auswahl, Einordnung, Ton, Quellenbewertung und Veröffentlichung redaktionell bei Benjamin Metzig verantwortet bleiben. Mehr zum Profil: Autorenprofil von Benjamin Metzig.
















































































Kommentare