
Der Atem der Erde beginnt im Mangankern: Wie Photosystem II Wasser zur Elektronenquelle macht
- Benjamin Metzig
- 3. Juni
- 6 Min. Lesezeit

Fast jeder Atemzug setzt eine biochemische Vorarbeit voraus, die erstaunlich unerquicklich beginnt: Jemand muss Wasser Elektronen entreißen. Nicht Zucker, nicht Kohlendioxid, nicht "die Pflanze" im Allgemeinen, sondern ein eng gebauter Proteinkomplex in der Thylakoidmembran erledigt den riskantesten Teil der oxygenischen Photosynthese.
Dieser Komplex heißt Photosystem II. Er ist keine grüne Deko im Lehrbuch, sondern ein Apparat, der Lichtblitze in oxidierende Gewalt übersetzt, Elektronen aus Wasser zieht und dabei den Sauerstoff freisetzt, an dem komplexes Leben hängt. Je genauer man ihn anschaut, desto weniger wirkt er wie ein stilles Naturwunder und desto mehr wie eine hochbelastete Maschine, die nur deshalb funktioniert, weil sie Zeit, Metalle, Wasserwege und Reparaturzyklen zugleich organisiert.
Kernaussagen
Photosystem II ist der biochemische Startpunkt der oxygenischen Photosynthese: Hier wird Wasser zum Elektronendonor, und hier entsteht der molekulare Sauerstoff.
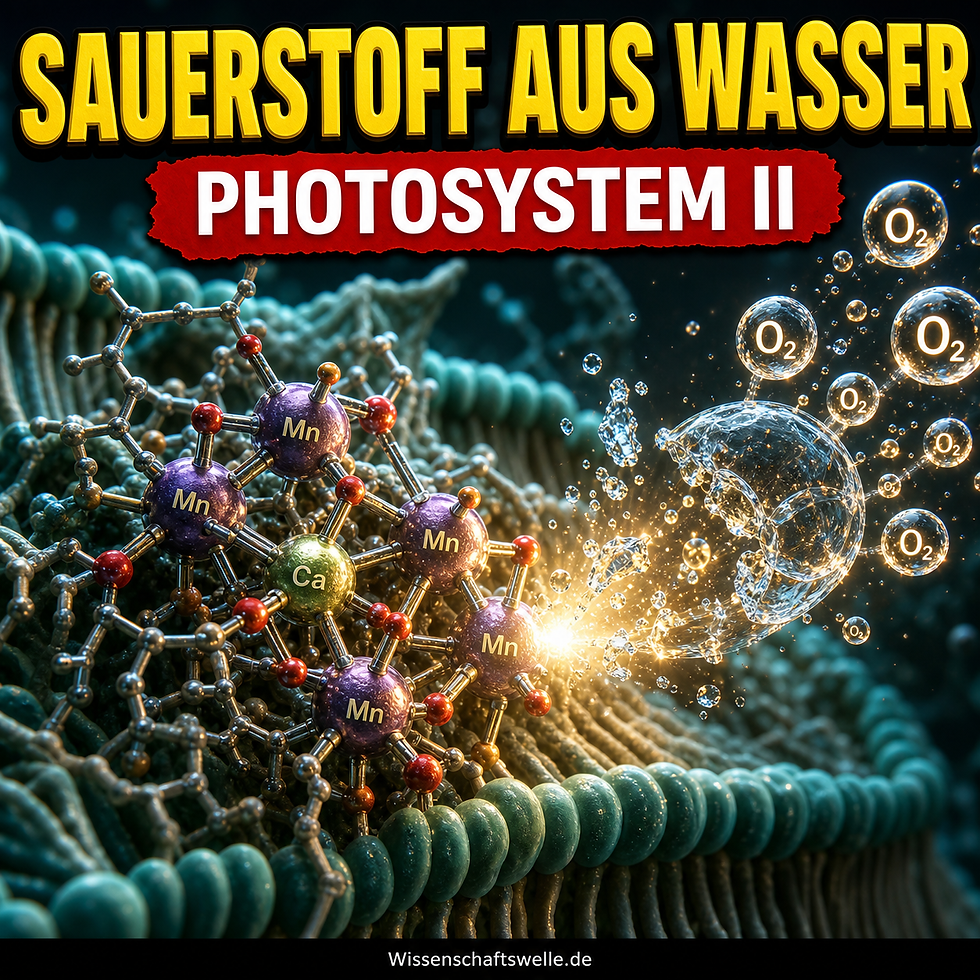
Die eigentliche Schwerarbeit leistet ein Mn4CaO5-Cluster, das vier Oxidationsäquivalente zwischenspeichert, bevor O2 gebildet werden kann.
Licht reicht nicht als einzelner "Schalter". Erst der gestufte Kok-Zyklus macht aus vier separaten Photonen eine kontrollierte Mehr-Elektronen-Chemie.
Neue Strukturarbeiten zeigen immer präziser, wie Wasser und Protonen den Katalysekern erreichen oder verlassen; die letzte O-O-Bindung ist trotzdem noch kein vollständig abgehaktes Kapitel.
Diese Reaktion treibt nicht nur Pflanzen an. Sie war eine Voraussetzung für die oxygenierte Atmosphäre und damit für die spätere Geschichte komplexen Lebens.
Wasser ist als Rohstoff fast beleidigend unkooperativ
Wer Elektronen aus Wasser gewinnen will, sucht sich keinen bequemen Partner. Wasser ist chemisch stabil, in Zellen allgegenwärtig und als Elektronendonor unerquicklich. Genau darin liegt die Zumutung des Photosystem II: Es muss einen Stoff oxidieren, den andere biologische Reaktionen meist nur als Lösungsmittel oder Transportmedium behandeln.
Die Netto-Reaktion ist knapp formuliert: Aus zwei Wassermolekülen werden ein O2-Molekül, vier Protonen und vier Elektronen. In der Praxis ist das keine einzelne Reaktion, sondern eine Folge präzise getakteter Zwischenzustände. Das große Problem ist nicht nur, Elektronen zu entfernen. Es geht auch darum, positive Ladung zwischenzuspeichern, Protonen rechtzeitig loszuwerden und zwei Sauerstoffatome in genau dem Moment aneinanderzubringen, in dem die Chemie das zulässt.
Aus einem Lichtblitz wird ein Elektronenloch
Photosystem II sitzt am Anfang der lichtabhängigen Reaktionskette. Antennenpigmente sammeln Photonen ein und leiten die Energie zum Reaktionszentrum weiter, bis das Spezialchlorophyll P680 in einen angeregten Zustand gerät. Dann passiert der entscheidende Bruch: Ein Elektron springt auf die Akzeptorseite und läuft über Pheophytin und gebundene Quinone weiter in die Elektronentransportkette. Was zurückbleibt, ist P680+, ein außergewöhnlich starkes Oxidationsmittel.
Diese positive Ladung ist der eigentliche Motor der Wasserspaltung. Sie wird nicht direkt an Wasser angelegt, sondern über eine redoxaktive Tyrosin-Seitenkette, meist YZ genannt, zum katalytischen Metallkern weitergereicht. So wird das Reaktionszentrum von der brutalen Chemie ein Stück weit entkoppelt, ohne dass die Oxidationskraft verloren geht. Eine gute Übersicht darüber, wie diese gekoppelte Ladungsverschiebung das System organisiert, liefert das Review Water oxidation in photosystem II.
An dieser Stelle wird auch klar, warum Eingriffe in Photosystem II so folgenreich sind. Wenn auf der Akzeptorseite der Elektronenfluss stockt, staut sich die ganze Reaktionslogik nach hinten auf. Genau diesen Zusammenhang sieht man auch im bereits veröffentlichten Beitrag Wo Herbizide Pflanzen treffen: Wie Chemie Elektronen, Pigmente und Enzyme lahmlegt, in dem die Verwundbarkeit des photosynthetischen Elektronentransports aus anderer Richtung sichtbar wird.
Der Mangankern speichert Zeit
Dass Photosystem II Wasser oxidieren kann, liegt nicht an einem magischen Einzelmolekül, sondern an einem Cluster aus vier Manganionen, einem Calciumion und fünf Brückensauerstoffen, kurz Mn4CaO5. Mangan ist hier nicht zufällig eingebaut: Es kann mehrere Oxidationsstufen stabil durchlaufen und eignet sich damit für das schrittweise Ansammeln oxidativer Äquivalente. Calcium trägt die Hauptredoxarbeit nicht selbst, stabilisiert aber Geometrie, Ligandenumgebung und das Wasserstoffbrückenmilieu des aktiven Zentrums mit. Die hochaufgelöste Struktur von Umena et al. 2011 machte diesen Kern erstmals so sichtbar, dass Liganden, gebundene Wassermoleküle und die geometrische Umgebung konkret diskutierbar wurden. Mit den femtosekundenscharfen XFEL-Daten von Suga et al. 2015 wurde das Bild robuster, weil sich strahlungsbedingte Strukturverzerrungen besser vermeiden ließen.
Der Kern funktioniert wie ein Speicher für vier oxidative "Schuldscheine". Jedes absorbierte Photon ermöglicht letztlich einen weiteren Oxidationsschritt am Metallcluster. In der klassischen Sprache des Kok-Zyklus läuft das System durch die S-Zustände S0 bis S4. Erst wenn genug Oxidationsäquivalente gesammelt wurden, kann die O-O-Bindung entstehen und Sauerstoff freigesetzt werden. Das ist der geniale Umweg des Systems: Nicht ein Photon muss eine Vier-Elektronen-Reaktion erledigen, sondern vier Lichtblitze laden nacheinander einen Katalysator auf, der die schwere Chemie zeitlich bündelt.
Die neueren Strukturserien von Li et al. 2024 zeigen, wie eng diese Aufladung mit kleinen Umlagerungen im Wasserumfeld des Clusters verknüpft ist. Photosystem II arbeitet also nicht nach dem Muster "Licht rein, Sauerstoff raus", sondern nach dem Muster "Lichtblitze einzeln einsammeln, Oxidationskraft zwischenlagern, Wasser präzise in Stellung bringen, dann erst die große Reaktion erlauben".
Wasser hinein, Protonen hinaus, alles im richtigen Takt
Sobald man akzeptiert, dass der Metallkern nicht isoliert arbeitet, wird das Thema noch interessanter. Der Katalyseort braucht Nachschub an Wassermolekülen und zugleich Wege, über die Protonen entweichen können. Beides darf nicht chaotisch passieren, weil sonst Ladungen, pH-Milieu und Wasserstoffbrücken-Netze nicht mehr zum Oxidationszustand des Kerns passen würden.
Genau hier sind die Strukturarbeiten der letzten Jahre besonders aufschlussreich. Hussein et al. 2021 verfolgten beim Übergang von S2 nach S3 Bewegungen in Wasser- und Protonenkanälen, die die funktionelle Arbeitsteilung solcher Pfade plausibler machen: Bestimmte Korridore scheinen eher Substratwasser zu liefern, andere eher Protonenabgabe zu erleichtern. Das heißt nicht, dass jetzt jede Route endgültig kartiert wäre. Aber die Vorstellung eines statischen Metallknotens in einem ansonsten passiven Protein ist damit endgültig zu klein.
Wer bei Wasserkanälen sofort an Transportproteine denkt, kann das mit dem älteren Wissenschaftswelle-Text über Aquaporine kontrastieren. Dort geht es um selektiven Durchlass durch Membrankanäle. Im Photosystem II dagegen geht es nicht primär um Durchsatz, sondern um katalytisch passend getimte Lieferung und Entlastung in unmittelbarer Nähe eines hochreaktiven Zentrums.
Der Sauerstoff fällt nicht einfach aus dem System
Die berühmte offene Frage lautet nicht mehr, ob Photosystem II Wasser spaltet. Das ist geklärt. Spannend bleibt, wie genau die O-O-Bindung im letzten Reaktionsabschnitt entsteht. Mehrere Modelle konkurrieren in Details, oft mit unterschiedlichen Rollen einzelner Sauerstoffatome oder Zwischenzustände. Gerade deshalb sind die jüngeren Momentaufnahmen aus serieller Kristallographie so wichtig.
Die Arbeit von Bhowmick et al. 2023 liefert strukturelle Hinweise auf Zwischenzustände während der O2-Bildung und des anschließenden Resets. Zusammen mit den S1-S2-S3-Daten aus 2024 verdichtet sich das Bild, dass zusätzliche Wassereinlagerung, Ligandenbewegung und Protonendynamik enger gekoppelt sind, als ältere Schemata es nahelegten. Viel spricht dafür, dass ein neu positioniertes Sauerstoffatom, oft als O6 diskutiert, in den kritischen Endschritt hineinspielt. Aber das ist keine Stelle, an der redaktionelle Sicherheit stärker sein sollte als die Literatur.
Genau darin liegt ein Reiz dieses Themas: Photosystem II ist eines der am besten untersuchten Enzymsysteme der Biochemie, und trotzdem bleibt die letzte Nanosekunde der Sauerstoffbildung wissenschaftlich lebendig. Das macht den Stoff interessanter als jede vereinfachte Formel von der "perfekten Naturmaschine".
Eine Hochleistungschemie, die regelmäßig neu aufgebaut werden muss
Es ist verführerisch, Photosystem II als makellos optimierten Apparat zu erzählen. Tatsächlich arbeitet der Komplex nahe an einer chemischen Schmerzgrenze. Die Oxidationskraft von P680+ ist so hoch, dass Fehlreaktionen und photochemische Schäden nie weit weg sind. Pflanzen, Algen und Cyanobakterien betreiben deshalb nicht nur Lichtchemie, sondern auch einen ständigen Wartungsbetrieb, bei dem insbesondere Komponenten des Reaktionszentrums immer wieder repariert oder ersetzt werden.
Das ist kein Randdetail, sondern Teil der eigentlichen Pointe. Photosystem II ist nicht deshalb beeindruckend, weil es mühelos Sauerstoff erzeugt. Es ist beeindruckend, weil es diese aggressive Chemie in einem biologischen Milieu überhaupt beherrschbar macht. Wer nur das Endprodukt O2 betrachtet, verpasst die eigentliche Leistung: kontrollierte Instabilität.
Warum diese Reaktion die Erdgeschichte umgeschrieben hat
Die große historische Bedeutung liegt nicht darin, dass moderne Blätter hübsch grün sind. Oxygenische Photosynthese, wie wir sie heute mit Photosystem II kennen, verschob die Chemie des Planeten. Der Review von Fischer et al. 2016 ordnet ein, wie die Evolution dieser Wasseroxidation mit der späteren Oxygenierung von Ozeanen und Atmosphäre verknüpft ist. Der Sauerstoffgehalt der Erde stieg nicht in einem einzigen harmonischen Sprung, aber ohne diese biochemische Innovation gäbe es weder eine ozonreiche Schutzschicht noch dieselben energetischen Möglichkeiten für aeroben Stoffwechsel.
Gerade deshalb lohnt es sich, Photosystem II nicht als botanisches Detail abzuheften. Es ist ein planetarer Schwellenmechanismus. Die Biologie hat hier nicht bloß Licht "genutzt", sondern einen Weg gefunden, das häufigste geeignete Reduktionsmittel der Erdoberfläche, Wasser, in einen global wirksamen Elektronenspender zu verwandeln.
Wer die Brücke zur Technik ziehen will, findet im Beitrag über Photokatalyse einen sinnvollen Kontrast. Technische Systeme versuchen ebenfalls, Licht in chemische Arbeit zu übersetzen. Aber der Abstand zur biologischen Eleganz und Robustheit von Photosystem II ist immer noch groß, gerade weil hier Mehr-Elektronen-Chemie, Protonenmanagement, Selbstschutz und Reparatur zusammenspielen.
Was man sich von Photosystem II merken sollte
Photosystem II zeigt, dass die vielleicht folgenreichste Reaktion der Biosphäre nicht aus einer großen Geste besteht, sondern aus sauber getakteter Vorarbeit. Vier Photonen, ein hochreaktiver Chlorophyllkern, eine Tyrosin-Brücke, ein Mangankomplex, zwei Wassermoleküle, mehrere Protonenwege und ein eng begrenzter Moment, in dem Sauerstoff entstehen darf. Das ist kein beiläufiger Nebeneffekt des Pflanzenlebens. Es ist die aufwendige biochemische Vorbedingung dafür.
Wenn man diese Reaktion kleinredet, klingt Sauerstoff wie etwas, das Blätter eben "mitmachen". Schaut man genauer hin, wirkt derselbe Sauerstoff eher wie das Produkt einer kontrollierten Zwangsarbeit am Wasser. Und genau das macht Photosystem II so bemerkenswert: Der Atem der Erde beginnt nicht im großen Ganzen, sondern in einem Metallkern, der Licht in Mangel übersetzt und daraus Lebenstauglichkeit macht.
Autorenprofil
Benjamin Metzig ist Gründer, Autor und redaktionell Verantwortlicher von Wissenschaftswelle.de. Wissenschaftswelle ist ein persönlich geführtes redaktionelles Wissensprojekt, das komplexe Themen aus unterschiedlichen Fachbereichen sorgfältig recherchiert, strukturiert und verständlich aufbereitet. Moderne Recherche-, Analyse- und KI-Werkzeuge dienen dabei als Unterstützung, während Auswahl, Einordnung, Ton, Quellenbewertung und Veröffentlichung redaktionell bei Benjamin Metzig verantwortet bleiben. Mehr zum Profil: Autorenprofil von Benjamin Metzig.
















































































Kommentare